艾弗里利用减法原理严谨控制自变量,想要证明DNA是遗传物质。然而他的论文发表后却引发轩然大波,备受质疑。原因主要有两点:一是当时人们受主流思想限制,不愿接受这个结论;二是受科技水平限制,艾弗里没有将蛋白质和DNA彻底分开,无法直接单独观察二者的作用。
我们的实验思路没有变,依然要把蛋白质和DNA完全分开,既然现有技术做不到,就只能另辟蹊径。
孟德尔曾经说过:任何实验的价值和效用,取决于实验材料对实验目的的适合性。我们需要一种更简单的生物,最好只由蛋白质和DNA构成,且蛋白质和DNA能自然分离。
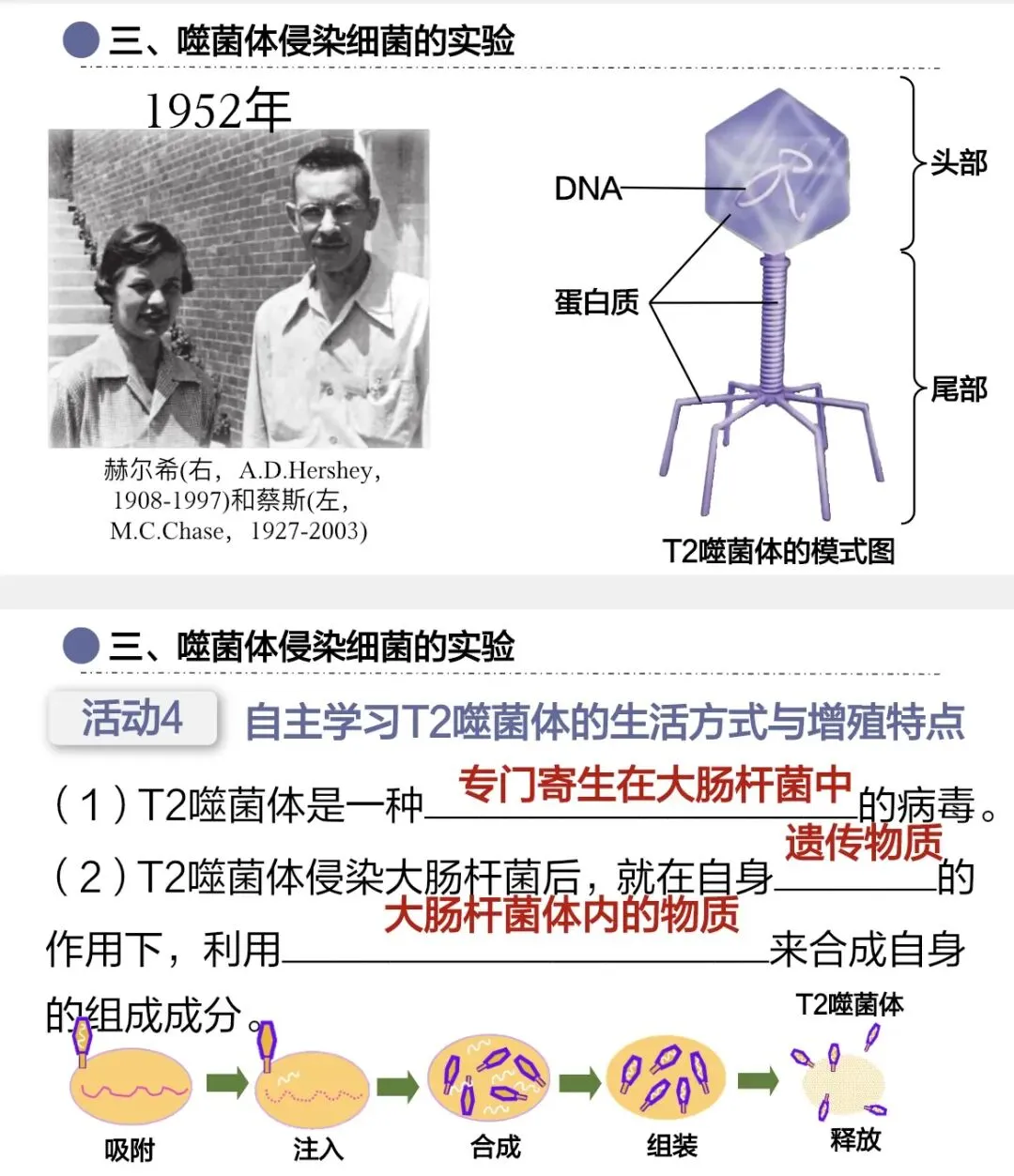
大家想到了,这就是病毒。1952年,赫尔希和蔡斯选择了理想的实验材料——T2噬菌体。
T2噬菌体结构非常简单,只有蛋白质外壳和内部的DNA。它是专一寄生在大肠杆菌体内的病毒,会先吸附到大肠杆菌表面,再进行增殖。侵染大肠杆菌后会产生大量子代噬菌体,这一定是遗传物质注入大肠杆菌后,控制增殖的结果。
我们只需要追踪:谁进入了大肠杆菌,谁出现在子代中,谁就是遗传物质。
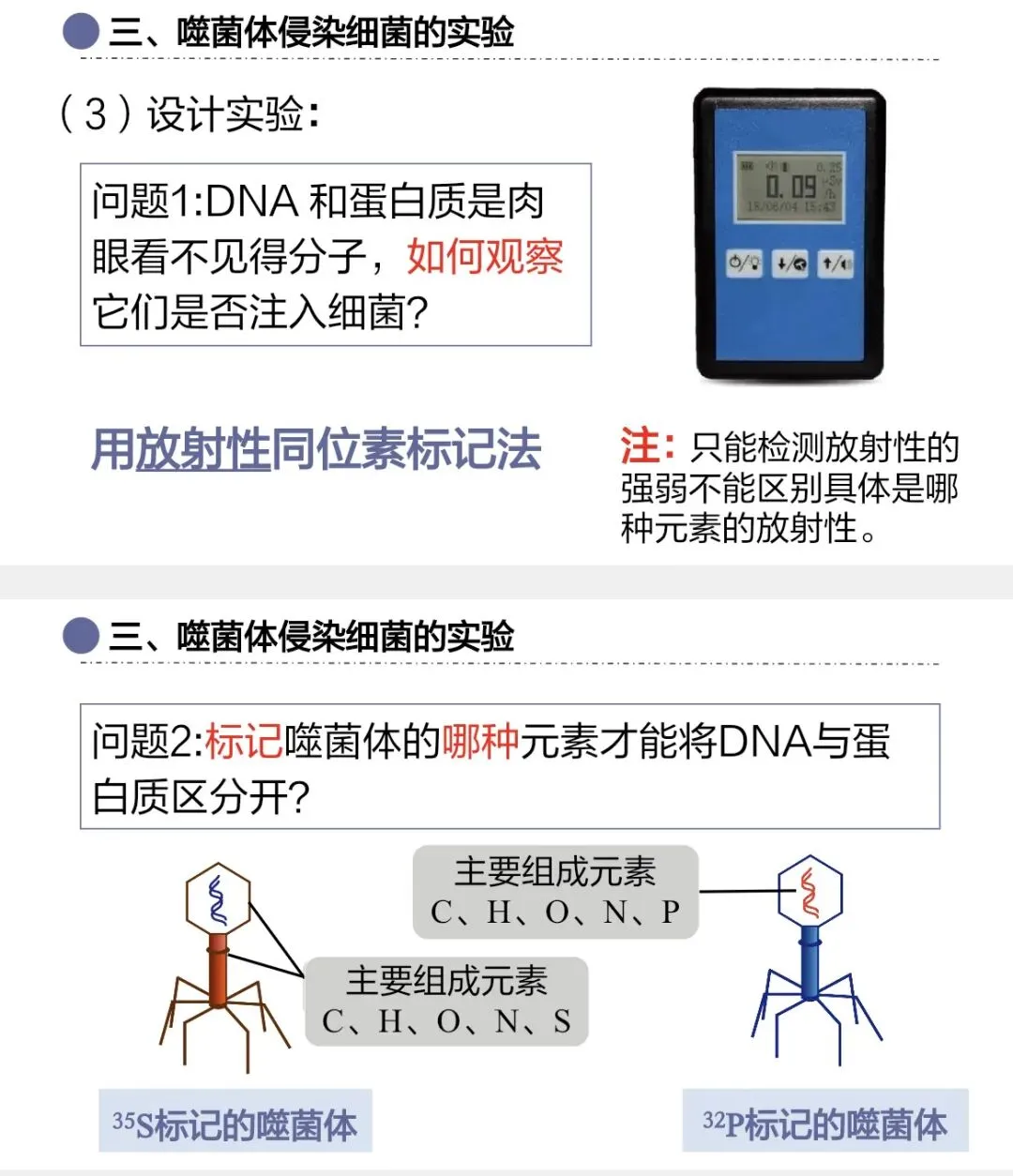
大家在设计实验时会遇到技术难题:蛋白质和DNA的分子很小,怎么追踪它们的行为?用放射性同位素标记法。
选择哪种同位素分别标记蛋白质和DNA?用³⁵S标记蛋白质,用³²P标记DNA。为什么不选择C、H、O、N这些元素?因为二者都含有这些元素,无法区分。
如何让噬菌体带上放射性标记?
(学生回答)
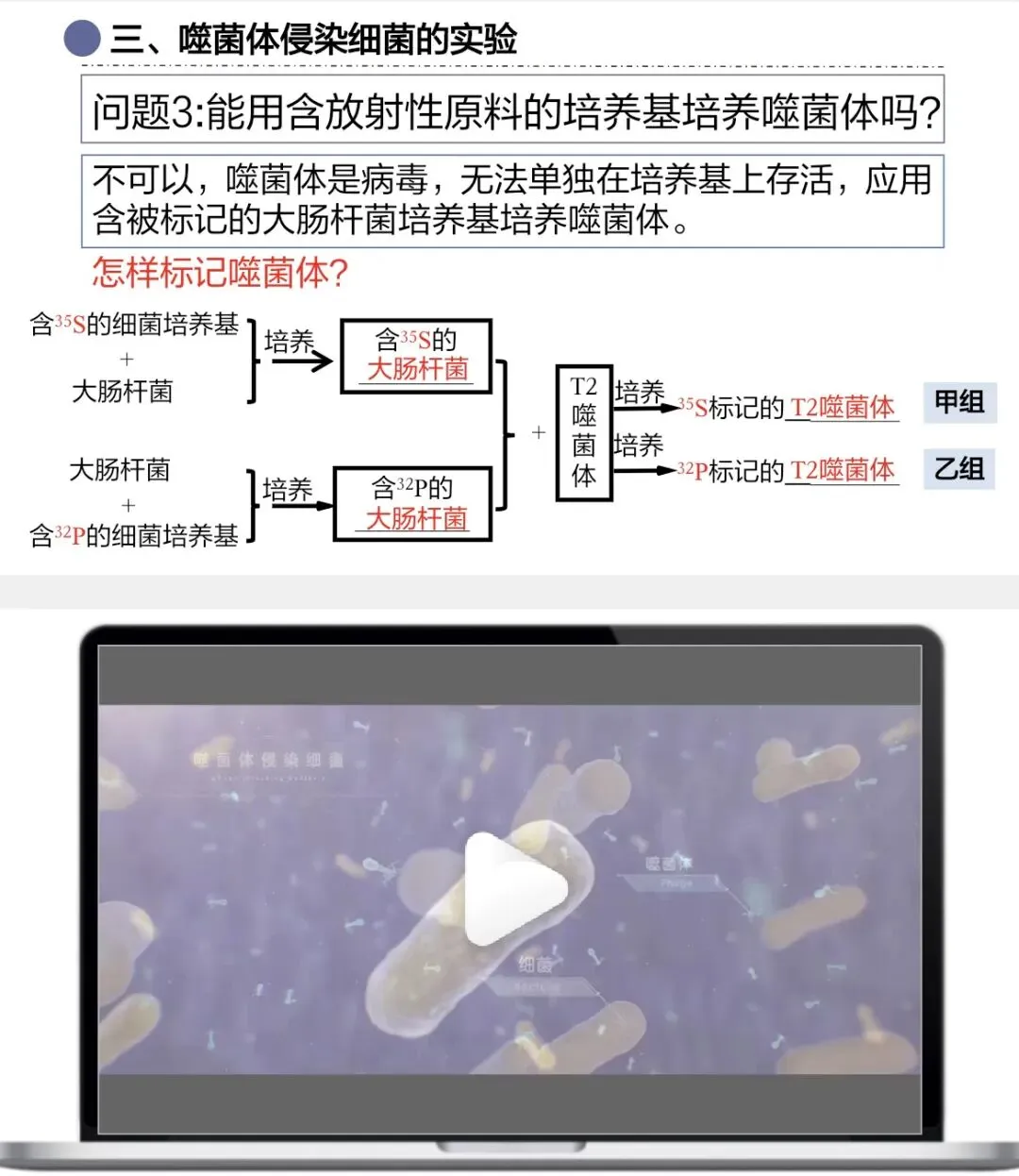
噬菌体是病毒,不能独立代谢,必须依赖活细胞,所以要先标记大肠杆菌,再用噬菌体去侵染标记后的大肠杆菌
标记时要分开做两组实验:一组标记蛋白质(³⁵S),一组标记DNA(³²P),不能同时标记,因为仪器只能检测放射性的有无和强弱,不能检测放射性种类。
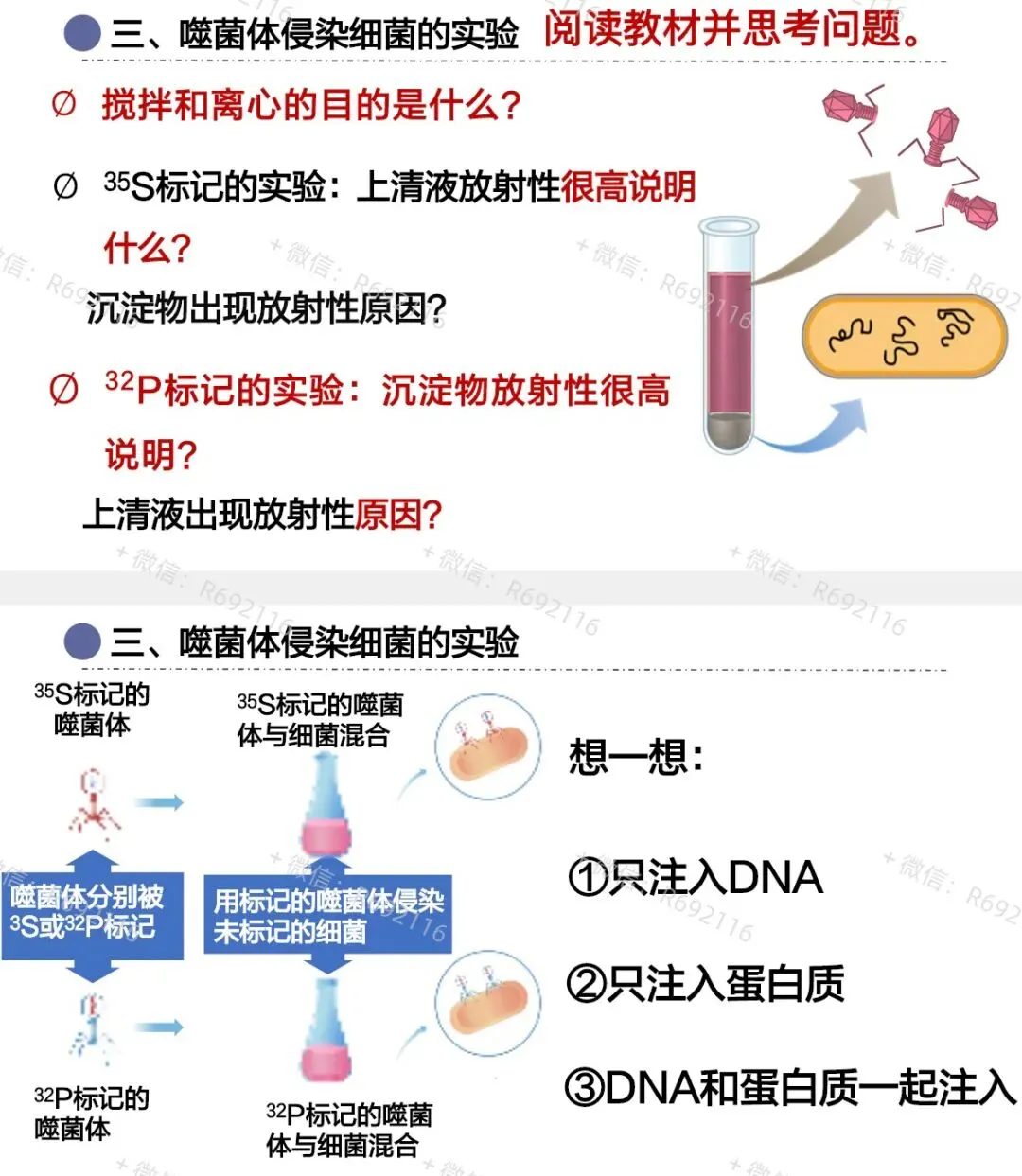
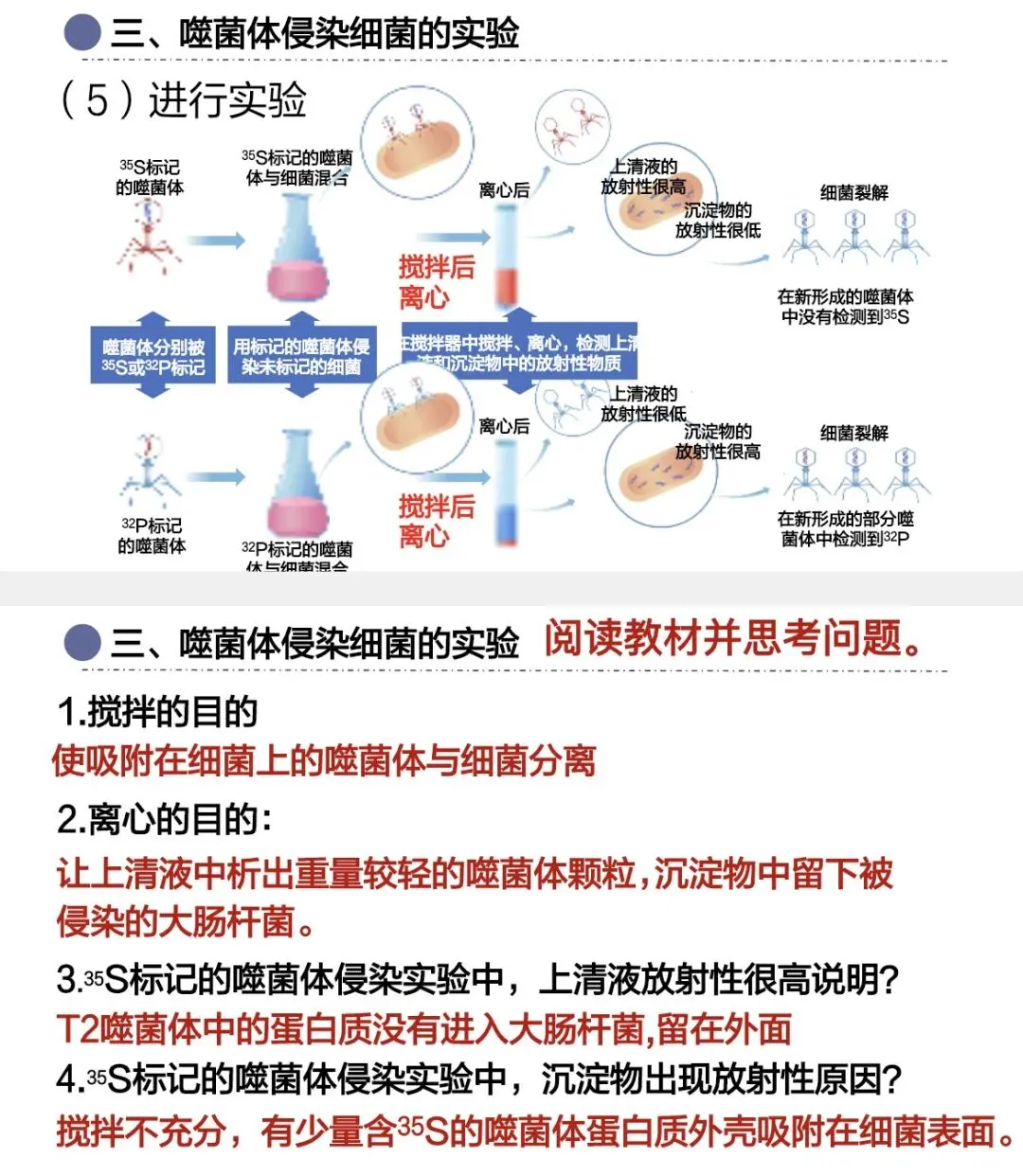
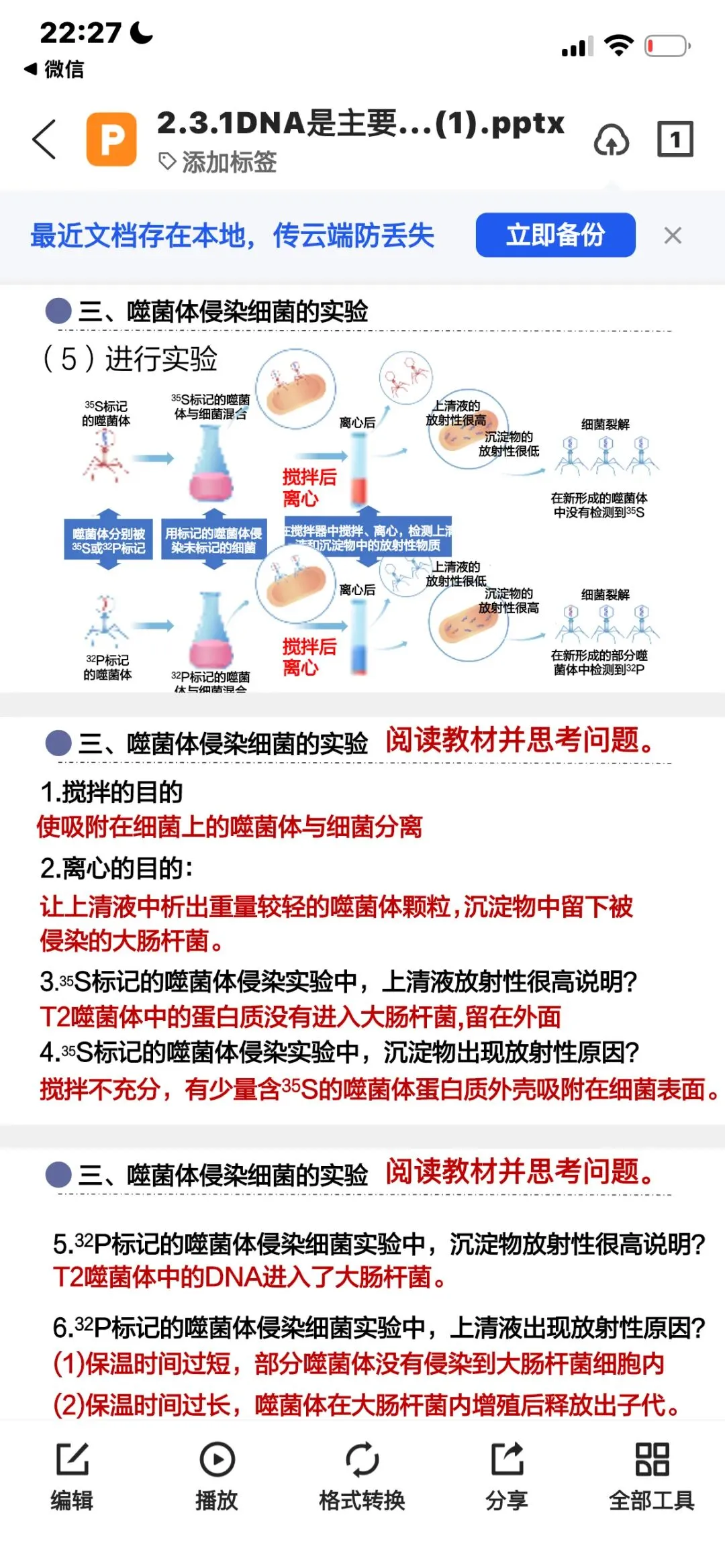
接下来让噬菌体侵染大肠杆菌:噬菌体先吸附在大肠杆菌表面,注入遗传物质。此时,带有放射性的蛋白质外壳还吸附在大肠杆菌表面,因此需要先搅拌,使蛋白质外壳与大肠杆菌分离,再离心。
离心后,较轻的噬菌体外壳分布在上清液,较重的大肠杆菌分布在沉淀物。最后检测离心管中放射性的位置即可。
实验结果:
³²P标记DNA组,放射性主要出现在沉淀物中;
³⁵S标记蛋白质组,放射性主要出现在上清液中。
这个结果说明:DNA进入大肠杆菌,蛋白质外壳没有进入。
作为遗传物质,还需要传递给子代。赫尔希和蔡斯进一步检测子代噬菌体,结果在子代中能检测到³²P标记的DNA,却检测不到³⁵S标记的蛋白质。这说明DNA传给了子代,DNA才是噬菌体的遗传物质。
是不是所有生物的遗传物质都是DNA?
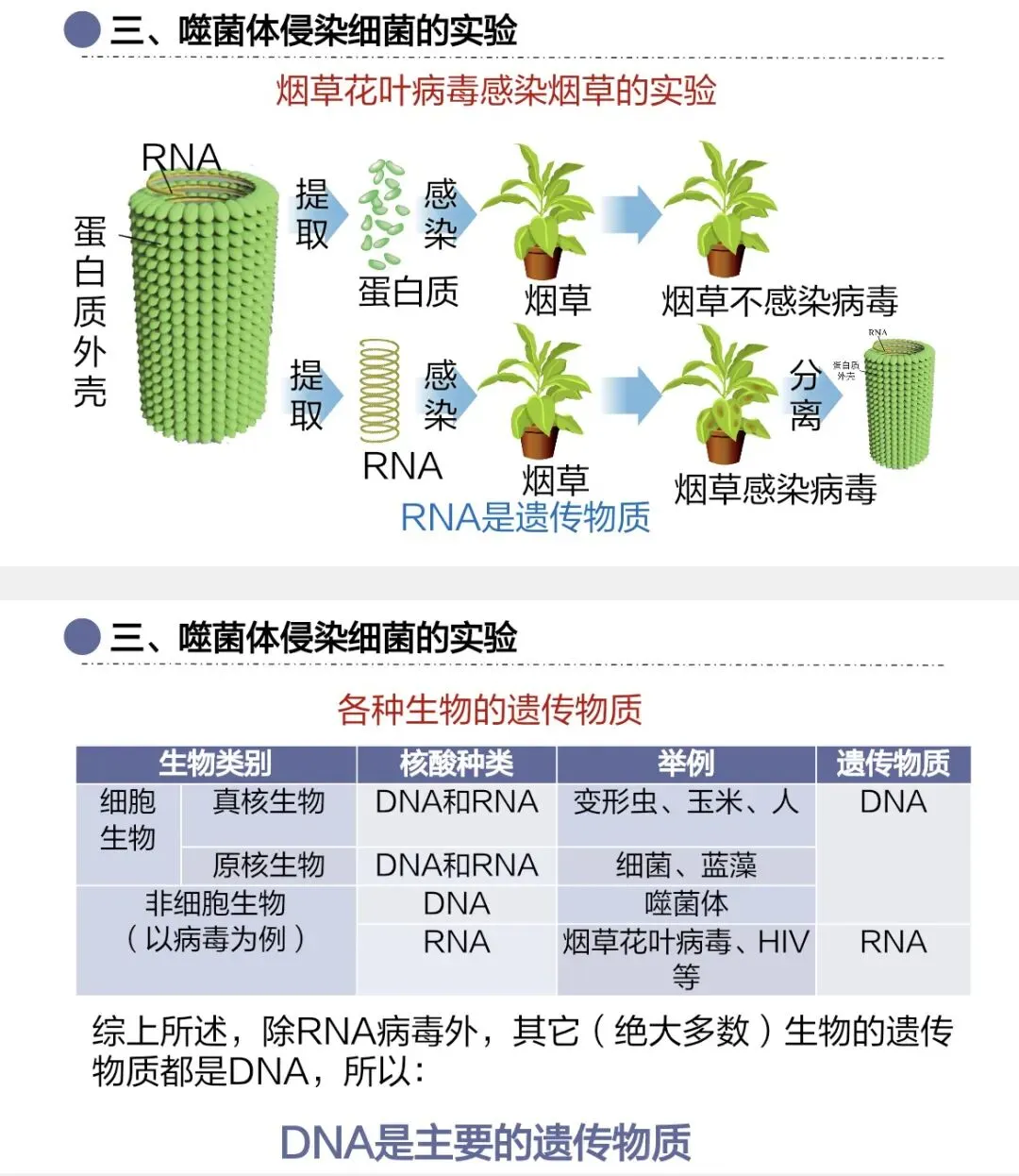
我们来看烟草花叶病毒,它只由蛋白质和RNA构成。它的遗传物质是什么?该如何证明?
(学生回答)
可以用类似的方法,单独研究蛋白质和RNA的作用,也可以用减法原理:去除蛋白质或RNA,观察能否增殖。
实验结论:烟草花叶病毒的遗传物质是RNA。
由此可知,并非所有生物的遗传物质都是DNA。绝大多数生物的遗传物质是DNA,少数生物的遗传物质是RNA,因此我们认为DNA是主要的遗传物质。
现在我们回顾今天的探索历程:从20世纪初到1952年,人们的观点从推测蛋白质是遗传物质,到证明DNA是遗传物质,再到认同DNA是主要的遗传物质,前后经历了半个多世纪。
科学理论的构建不是一帆风顺的,是不断被推翻、重建、再完善的过程。这个过程离不开实验材料的不断更新,更离不开技术水平的不断进步。科学与技术相互支持、相互促进、共同发展。
我们对遗传物质的探索绝不是终点,而是新的起点,因为科学无止境,探索不停息。
剩下的时间,请大家完成课后习题。
好,同学们,下课。